Vanskelige overganger
(Fra boka 'The Design of Life' kap 1,6 og 7; av W. Dembski og J. Wells)
Tematikken i det følgende er hvilke 'gap' som måtte fylles av evolusjonære mekanismer, dersom de alene skulle forklare virkeligheten slik den framtrer i dag. De er bare ment som eks, ikke på noen måte som utfyllende forklaringer. Grunnen til at de er valgt ut, er både fordi de peker på vanskelige utfordringer -og at de enten er lite kjent, eller ikke i tilstrekkelig grad tatt inn over seg av neo-darwinismen.
Bilde 1. Betydelige forskjeller når det kommer til detaljer
 I. Fysiske forskjeller mellom mennesker og sjimpanser
I. Fysiske forskjeller mellom mennesker og sjimpanser
I hvilken grad mennesker og sjimpanser er når vi ser på gener, har vi sett på tidligere -her. Her skal vi betrakte noe forskjeller i kroppsbygning og framtreden:
1. Sjimpansens føtter har gripeevne, i motsetning til menneskets.
2. Mennesker har fremstikkende nese, i motsetning til sjimpanser.
3. Hunkjønns-aper har ikke menstruasjonspause , i motsetning til hos mennesket.
4. Mennesket er eneste primat der hunnens bryster er tydelige, når de ikke dier.
5. Mennesker har et lag med fet underhud, i likhet med hvaler og flodhester, i motsetning til hos aper.
6. Han-aper har et 1cm ben i penis, i motsetning til hos mennesket.
7. Mennesker er for det meste høyre-hendte. Sjimpanser benytter begge hender uten preferanser.
8. Mennesker svetter, i motsetning til aper.
9. Mennesker kan bevisst holde pusten, i motsetning til aper.
10 Mennesker er de eneste primater som gråter.
Dette er noen få av flere tydelige fysiske forskjeller mellom mennesker og sjimpanser. Men hovedforskjellen sitter i intellektuelle, lingvistiske og moralske evner hos mennesket.
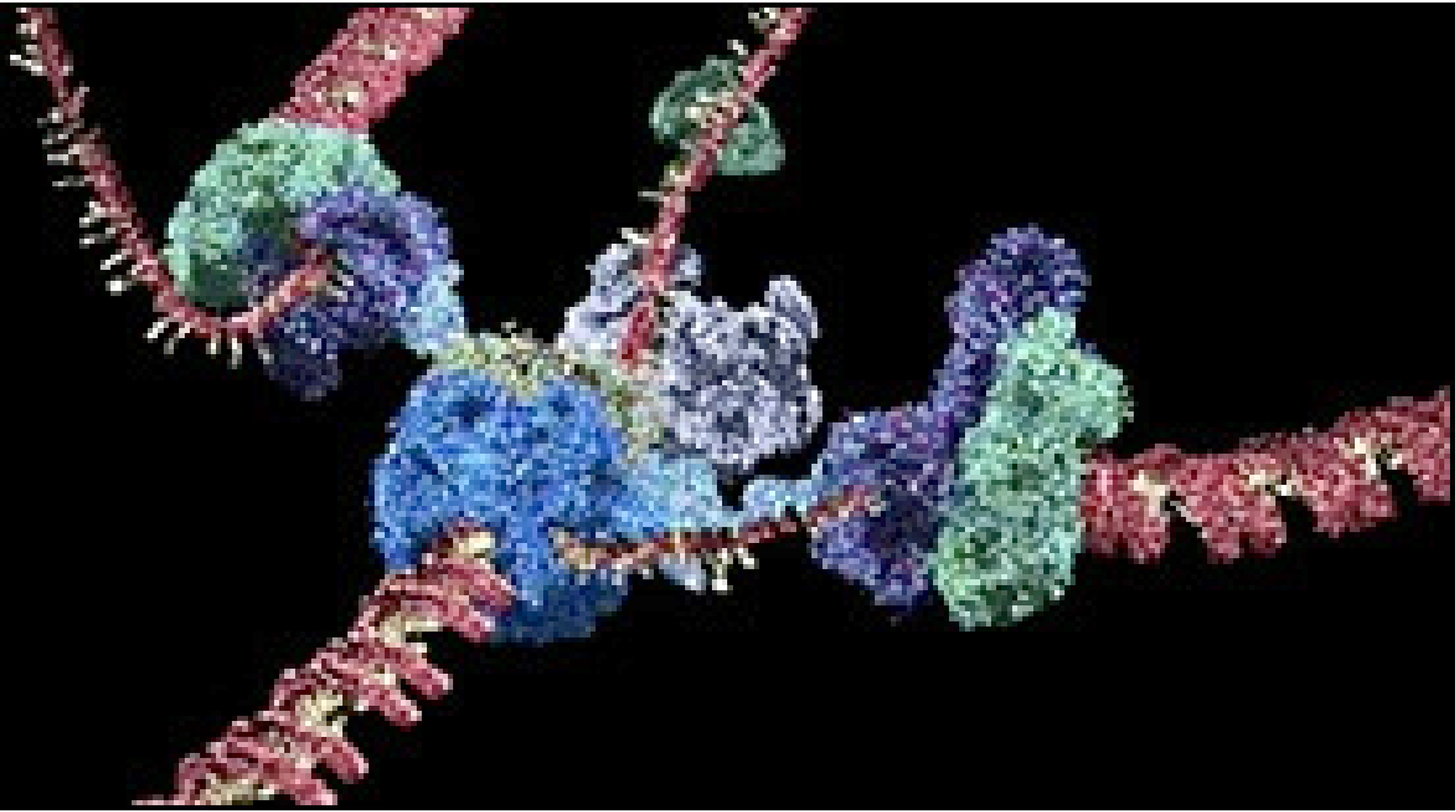 II. Molekylære maskiner
II. Molekylære maskiner
Biologi-journalen Cell utga i februar 1998 en spesialutgave knyttet til 'makro-molekylære maskiner'. Alle celler benytter komplekse molekylære maskiner til prosessere informasjon, konvertere energi, metabolisere næringsstoffer, bygge proteiner og transportere materiale gjennom membraner. Bruce Alberts fhv. president i National Academy of Sciences, introduserte emnet med en artikkel som het: "The Cell as a Collection of Protein Machines." Der skrev han: " Vi har alltid undervurdert celler. . Hele cellen kan sees som en fabrikk, som inneholder et utviklet nettverk av samvirkende produksjonslinjer, der hver er bygd opp av et sett store protein-maskiner .. lik maskiner oppfunnet av mennesker for å håndtere den makroskopiske verden, inneholder disse protein-sammenkoblingene høyst koordinerte bevegelige deler. (1)
Bilde 2. DNA-polymerase-maskin
 Nesten 6 år senere (Des. 2003), publiserte BioEssays sin egen spesialutgave av 'molekylære maskiner'. I introduksjonen til denne utgaven, bemerket redaktøren i BioEssays, Adam Wilkins, Artiklene i denne utgaven demonstrerer noen slående paralleller mellom tilvirkede og biologiske/molekylære maskiner. i) Molekylære maskiner, lik menneskelig tilvirkede maskiner utfører høyst spesifikke funksjoner. ii) Molekylære maskinkomplekser omfatter multiple deler som samvirker i distinkte og presise måter, med definerte input og output-funksjoner. iii) Mange av disse maskinene har deler som kan bli benyttet i andre molekylære maskiner ( i det minste med små modifikasjoner), sammenlignet med utbyttbare deler i kunstige maskiner. Sist men ikke minst, har de hoved-kjennetegnet på maskiner: de omgjør alle energi til en form for 'arbeid'. (2)
Nesten 6 år senere (Des. 2003), publiserte BioEssays sin egen spesialutgave av 'molekylære maskiner'. I introduksjonen til denne utgaven, bemerket redaktøren i BioEssays, Adam Wilkins, Artiklene i denne utgaven demonstrerer noen slående paralleller mellom tilvirkede og biologiske/molekylære maskiner. i) Molekylære maskiner, lik menneskelig tilvirkede maskiner utfører høyst spesifikke funksjoner. ii) Molekylære maskinkomplekser omfatter multiple deler som samvirker i distinkte og presise måter, med definerte input og output-funksjoner. iii) Mange av disse maskinene har deler som kan bli benyttet i andre molekylære maskiner ( i det minste med små modifikasjoner), sammenlignet med utbyttbare deler i kunstige maskiner. Sist men ikke minst, har de hoved-kjennetegnet på maskiner: de omgjør alle energi til en form for 'arbeid'. (2)
Selv om de nevnte forfattere (Alberts og Wilkins) er neo-darwinister og ser cellens fantastiske kompleksitet som produkt av darwinistisk evolusjon, og således bare som tilsynelatende designet. I 1990 årene begynte imidlertid forskere å utfordre det neo-darwinistiske synet og argumentere at slike molekylære maskiner utviser evidens på design. En som ledet denne an i denne utviklingen var Michael Behe.
III. Ikke-reduserbare systemer.
I 1996 publiserte Michael Behe en bok som het Darwins Black Box. I denne boka beskrev han hvordan neo-darwinismen feilet i å forklare opprinnelsen til komplekse, molekylære maskiner inni cellen. Men han stoppet ikke der, han argumenterte også for at disse molekylære maskiner oppsto ved faktisk design. Sentralt for argumentet hans var ideen om ikke-reduserbar kompleksitet. Et funksjonelt system er ikke-reduserbart komplekst, om det inneholder et samvirke av flere deler, som ikke kan forenkles uten å ødelegge systemets basisfunksjon. Vi kaller dette flerdelte undersystemet for systemets ikke-reduserbare kjerne. (2)
Å si at et system er ikke-reduserbart innebærer å si at ikke noe system med enklere kjerne kan utføre systemets basis-funksjon.
Denne består av 3 ting:
1) Det som systemet utfører i sin naturlige kontekst: dette er kjent som systemets basisfunksjon (eller hovedfunksjon).
2. Det minimale nivå av funksjonalitet som behøves for at systemet skal fungere tilstrekkelig i sin naturlige kontekst (systemets minimumsfunksjon).
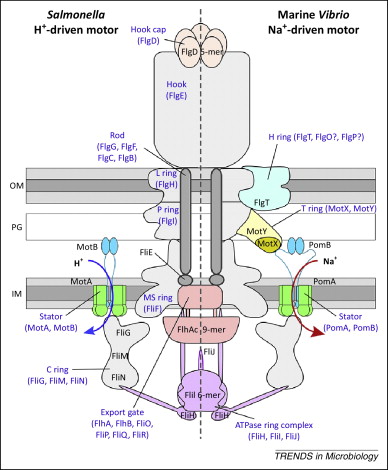
3. Måten som systemet utfører sin hovedfunksjon på (systemets funksjonsmodus). Fordi dette siste hører med, er også midlene som benyttes inkludert (f.eks. utfører lim og spiker noe av samme funksjon, men måten det skjer på er helt forskjellige.) Biologien er stappfull av ikke-reduserbare komplekse protein-maskiner. Faktisk er det vist at de fleste proteiner arbeider sammen med multiple proteiner for å utføre cellulære funksjoner. Således tilhører de fleste proteiner funksjonelt integrerte systemer. Av disse er mange ikke-reduserbart komplekse. Michael Behe nevner f.eks. blod-koagulerings-systemet, cilia og bakterie-flagellen. Flagellens intrikate maskineri krever koordinert interaksjon av omkring 30 proteiner og i tillegg ca. 20 for å assistere denne. Likevel er fraværet fra hvilken som helst av dem, tilstrekkelig til at fullstendig tap av motor-funksjon. (3) Disse proteinene former de ikke-reduserbare deler av flagellen.
Bilde 3. Funksjons-skisse for enkeltdeler i bakterieflagellen.
Bilde 4. Bakterie-flagell (skjematisk)

Mulige forklaringer
For å forklare ikke-reduserbar kompleksitet, så henviser neo-darwinister alltid til indirekte utviklings-stier. Det innebærer at ikke bare utvikler en ny struktur seg, men også dens tilknyttede funksjon. Til sammenligning er en direkte utviklings-sti, så utvikler og forbedrer naturlig seleksjon en struktur som allerede tjener en gitt funksjon, som ikke endrer seg. Siden funksjonen til et ikke-reduserbart komplekst system ikke er oppnådd før siste del er på plass, så måtte et slikt system bli produsert i ett nå. Og det er absurd at blinde, ikke-styrte krefter kan avstedkomme dette. Dermed blir det kun gjennom indirekte utviklings-stier at steg-for-steg oppbygning kan finne sted.
Det har vært forsøkt noen abstrakte scenarier for hvordan ikke-reduserbar kompleksitet kan oppstå. F.eks. har Allen Orr skissert denne muligheten: Et ikke-reduserbart system kan bygges gradvis ved å tilføye deler, som initielt er fordelaktige, men etterhvert blir uunnværlige. Logikken går som følger: Sett at en del A utfører en jobb -mindre godt riktignok. Del B tilføyes fordi den hjelper A. Denne nye delen er ikke essensiell, men forbedrer kun virkemåten. Men senere kan kanskje A eller en annen del endre seg på et slikt vis at B nå blir uunnværlig. Denne prosessen fortsetter i det flere deler blir føyd til systemet. Til slutt kan mange deler være uunnværlige. (4)
Det er uklart hvilke funksjoner eller jobber, som Orr her refererer til. Åpenbart må funksjonene underveis være ulike den endelige funksjonen, fordi den endelige funksjonen blir utført av et ikke-reduserbart system, og ikke kan utføres av noe annet system med en betydelig forenklet reduserbar kjerne. Men så kommer vi opp i samme problem som tidligere: Det er ikke noe empirisk bevis at ikke-reduserbare systemer blir til ved denne måten å tilføye komponenter på, og så etter hvert gjøre dem uunnværlige. Det som må kreves for å gjelde som vitenskapelige forklaringer, er detaljerte, testbare konstruksjoner eller modeller som viser hvordan neo-darwinistiske mekanismer kan produsere ikke-reduserbare komplekser som bakterieflagellen. Det eneste som har vært anført, som kan ligne en slik forløper', men som antagelig viser seg å være en etterfølger til bakterieflagellen, er TTSS -så henviser til tidligere omtale av denne -lenke.
 Argumentet fra ikke-reduserbar kompleksitet
Argumentet fra ikke-reduserbar kompleksitet
I det store og hele har kritikere godtatt at Behe har sine biologiske fakta rette. De har også diskutert at neo-darwinistisk litteratur i det store og hele er taus om hvordan ikke-reduserbar kompleksitet opprinnelig kom til. De har bestridt Behes argument ut fra teoretiske og metodologiske grunner. Behes argument fra ikke-reduserbar kompleksitet foregir seg å vise at ikke-reduserbar kompleksitet er utenfor neo-darwinistiske mekanismer og at design forsyner en bedre forklaring av dem.
Argumentet fra ikke-reduserbar kompleksitet kan forstås ut fra tre nøkkelpunkter: a) et logisk b) et empirisk og c) et forklarende -poeng. Det logiske argumentet går som følger: Ikke-reduserbar kompleksitet er bevisbart utilgjengelig for direkte darwinistiske utviklingsveier. En direkte darwinistisk gangvei er en som utvikler et system ved at naturlig seleksjon inkrementelt forbedrer en gitt funksjon. I det systemet utvikler seg, utvikler ikke funksjonen seg, men forblir den samme. I dette tilfellet måtte systemet ha oppstått via utvikling av et enklere system, som utførte samme basis (hoved)-funksjon. Men fordi den ikke-reduserbare kjernen til et ikke-reduserbart komplekst system ikke kan forenkles uten å ødelegge basis-funksjonen, så kan ikke evolusjonære forløpere med enklere kjerne ha utført samme funksjon.
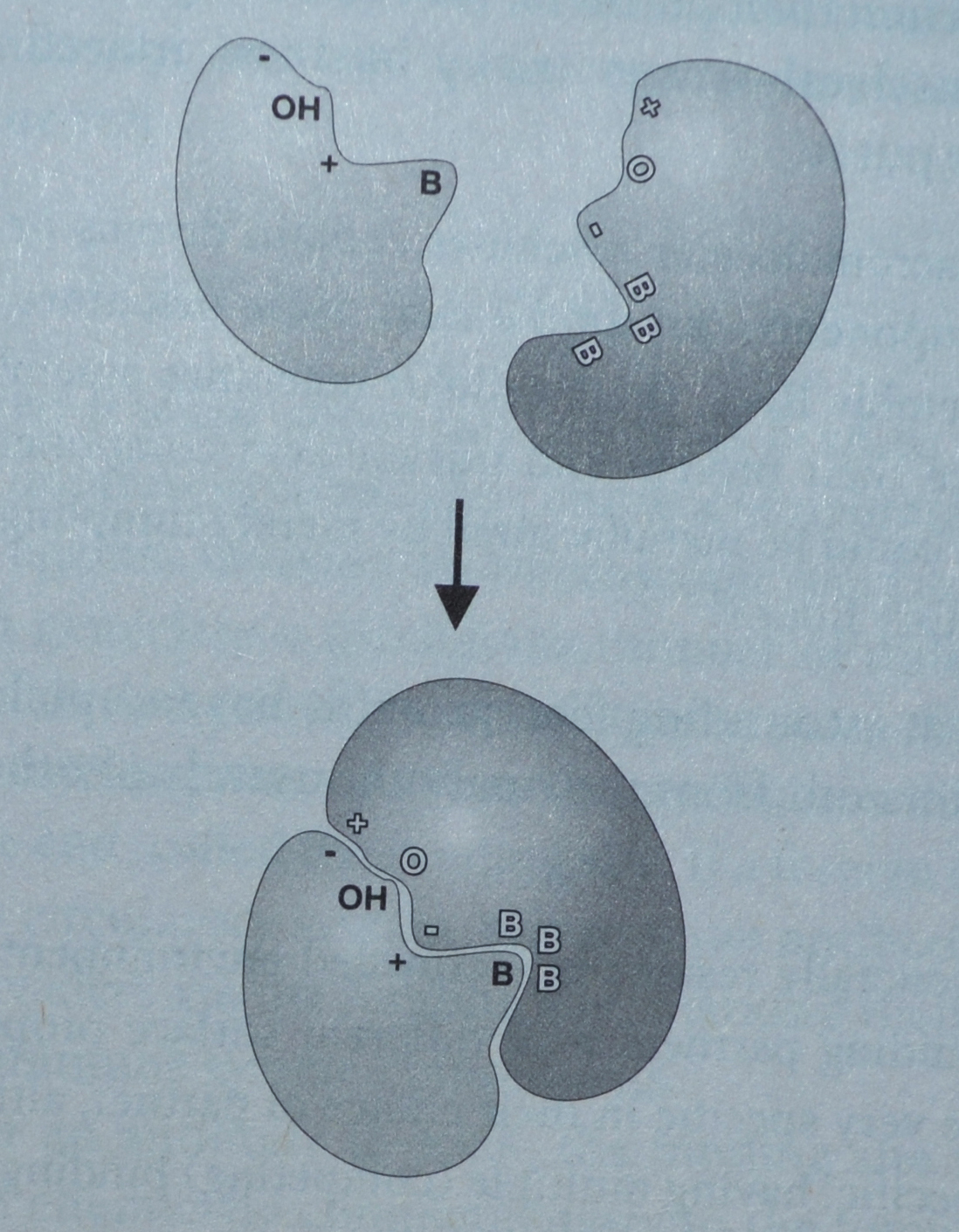
Bilde 5. Proteiner sammenkobles -både ved passform og ladning
Ved indirekte neo-darwinistiske utviklingsveier så samutvikler både struktur og funksjon seg. Tilfellet er at ingen indirekte neo-darwinistiske mekanismer er kjente for ikke-reduserbart komplekse systemer. Biologer har vært i stand til å identifisere undersystemer som utfører andre funksjoner. Men enhver rimelig komplisert maskin inkluderer subsystemer som kan utføre funksjoner atskilt fra den fullstendige maskinen. Det som trengs som vitenskapelig forklaring, er en sømløs neo-darwinistisk beretning som både er detaljert og falsifiserbar for hvordan undersystemer kan utsettes for ko-evolusjon og gradvis endres til et ikke-reduserbart komplekst system. Dette har som sagt manglet i den biologiske litteraturen, som i fall det skulle dukke opp, ville tilbakevise intelligent design.
Men hvordan kommer en fra det mislykkede i neo-darwinistiske mekanismer og over til at ID lykkes i så måte? Vitenskapelige forklaringer kommer i mange utgaver og fasonger, men den ene tingen de ikke kan klare seg uten, er årsaks-tilstrekkelighet (causal adequasy). En vitenskapelig forklaring trenger å påkalle en vitenskapelig forklaring som er tilstrekkelig til å fremkalle fenomenet som undersøkes. Ellers forblir fenomenet ikke-forklart. Emnet for undersøkelsen er ikke-reduserbar kompleksitet. Vi har sett at logiske forklaringer utelukker direkte neo-darwinistiske utviklingsveier. Empiriske forklaringer fjerner indirekte utviklingsstier, og heller ikke er det noen teori som kan forsyne bevis. Det foreligger altså ikke bevis for at natur-prosesser og lover er kausalt-tilstrekkelige til å forklare ikke-reduserbare systemer.
Naturlig seleksjon selekterer bare ut fra eksisterende funksjoner, så all vekt måtte bli på tilfeldige variasjoner å virkeliggjøre de riktige og nødvendige endringene. Det er imidlertid velkjent at intelligente årsaker kan forårsake dette, f.eks. ved at menneskelig intelligens kan forutse behov, planlegge, konstruere og implementere slik ikke-reduserbar kompleksitet. Darwin uttrykte det slik: "Dersom tjenlige variasjoner ikke inntreffer, så kan ikke naturlig seleksjon gjøre noe." (5) Likevel er aktuelle bevis for at tilfeldige variasjoner kan produsere suksessive endringer som trengs for å utvikle ikke-reduserbar kompleksitet lik null.
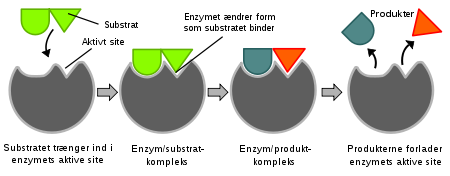
Bilde 6. Enzym med tilhørende substrat
 IV. Hindringer som evolusjon må overvinne
IV. Hindringer som evolusjon må overvinne
I forsøket på å koordinere de suksessive evolusjonære endringer som trengs for å frembringe ikke-reduserbare komplekse molekylære maskiner, møter de neo-darwinistiske mekanismene en rekke hindringer. Disse inkluderer følgende (6):
1. Tilgjengelighet. Er delene som trengs for å utvikle ikke-reduserbare, komplekse bio-kjemiske systemer slik som bakterieflagellen, en gang tilgjengelige? Sannsynlighet: Pt
2. Synkronisering. Er disse delene tilgjengelige til rett tid, slik at de kan inkorporeres, når de trengs i utviklende strukturer? Sannsynlighet: Ps
3. Lokalisering. Selv med deler som er tilgjengelige til trett tid til å inkluderes, så må delene frigjøres fra systemet de nå er integrert i (uten å skade disse systemene), og bli gjort tilgjengelige ved 'bygningsstedet' til det utviklende systemet. Sannsynlighet: 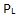
4. Forstyrrende kryss-reaksjoner. Gitt at de rette delene kan bli bragt sammen til rett tid på rett sted, hvordan kan feilaktige deler som ellers ville forpurre byggverket bli ekskludert fra 'bygningsstedet' for utviklende systemer? Sannsynlighet: 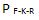
5. Grensesnitt-kompatibilitet. Er delene som rekrutteres for å inkluderes i et utviklende system gjensidig kompatible, slik at de kan sammenkobles tett, slik at når de en gang er sammenstilt at partene kan danne et varig fungerende system i livsløpet? Sannsynlighet: 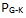
6. Orden-i-konstruksjonen. Selv med alle og bare korrekte deler som når rett plass til rett tid, og selv med fullt ut grensesnitt-kompatibilitet, vil de settes sammen i rett rekkefølge til å danne en fungerende enhet? Sannsynlighet: 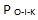
7. Konfigurasjon. Selv med alle de rette delene stilt opp til å settes sammen i rett rekkefølge, vil de arrangeres på rett måte for å danne et fungerende system? Sannsynlighet: Pk
Merk at hver av sannsynlighetene vi skrev opp, er avhengig av den foregående. F.eks. synkroniserings-sannsynligheten fastsetter sannsynligheten forutsatt at de nødvendige delene er tilgjengelige osv. Om vi kaller Opprinnelses-sannsynligheten for Po, så vil følgende ulikhet gjelde: Sannsynligheten til et utviklende neo-darwinistisk ikke-reduserbart komplekst system kan ikke overgå produktet av de ovenfor nevnte sannsynligheter:
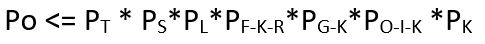
siden hver av sannsynlighetene underforstått vil være liten, vil det resulterende produktet bli desto mindre. Ingen av disse sannsynligheten må kalkuleres direkte, det holder å fastsette en øvre sannsynlighetsskranke for hver av dem. Dersom Opprinnelses sannsynligheten er liten, så vil det ikke-reduserbare komplekse systemet være både høyst usannsynlig og spesifisert, siden hver av disse systemene er spesifiserte i kraft av deres biologiske funksjon. Det følger at dersom opprinnelses-sannsynligheten er liten, så vil systemet utøve spesifisert kompleksitet, og siden spesifisert kompleksitet er en pålitelig empirisk markør for aktuell design, så følger at systemet i seg selv er designet.
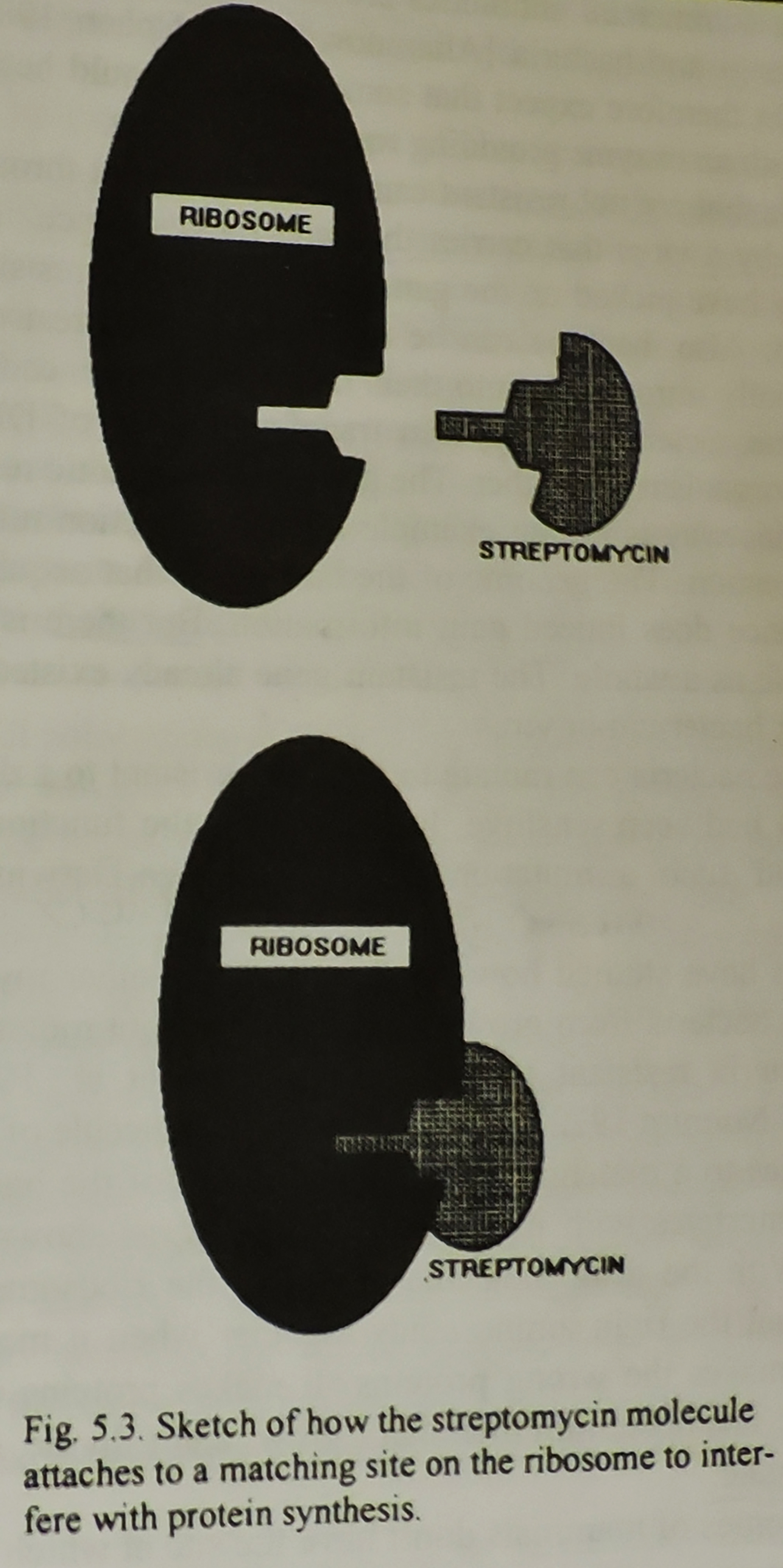
Bilde 7. Ribosom og streptomycin
Referanser:
1. Bruce Alberts, The Cell as a Collection of Protein Machines: Cell 9821 (8.febr, 1998): 291
2. Denne definisjonen er W. Dembskis refinement og generalisering av M. Behes originale definisjon.
3. Michael Behe, Darwins Black Box (New York: Free Press, 1996), 69-73
4. H. Allen Orr, "Darwin vs. Intelligent Design (Again)," boston Review (Des/Jan. 1996/1997): 29
5. Darwin, Origin of Species, 82.
6. Angus Menuge, Agents Under Fire: Materialism and the Rationality of Science (Lanham, Md: Rowman and Littlefield, 2004) ch. 4
Oversettelse og bilder ved Asbjørn E. Lund